 ISSN: 2822-0838 Online
ISSN: 2822-0838 Online

Hydro- and Osmo-priming Effects on Upland Rice Exposed to Drought Conditions at Vegetative and Reproductive Stages
Kartika Kartika, Benyamin Lakitan*, Rofiqoh Purnama RiaPublished Date : 2021-04-01
DOI : https://doi.org/10.12982/CMUJNS.2021.053
Journal Issues : Number 3, July-September 2021
Abstract Drought stress during vegetative and/or generative stages could cause massive reduction in rice yield. This study evaluated effectiveness of hydro- and osmo-priming on improving seed germination, growth and development of upland rice under drought stress during late vegetative, booting, or heading stage. Treatments consisted of hydro-priming and osmo-priming consisted of three polyethylene glycol (PEG) concentrations i.e. 10%, 15%, and 20%. Results showed that application of osmo-priming at 10% PEG required longer time (21.93 hours) to reach 50% germination, lower germination and lower vigor index. However, after germination, seeds primed with 10% PEG exhibited better seedling growth than other seed priming treatments. Effects of seed priming on yield components were overshadowed by drought exposures. Drought imposed during vegetative stage did not significantly affect yield; however, regardless of seed priming treatments, yield reduction was inevitable in rice plants exposed to drought during booting or heading stages. Rice plant shortened time to reached physiological maturity as an adaptive mechanism if drought treatment was applied during heading stage.
Keywords: Germination, Polyethylene glycol, Seedling growth, Seed priming, Yield component
Funding: This study was funded by Program Penelitian Unggulan Profesi Universitas Sriwijaya, Grant No. 0687/UN9/SK.BUK.KP/2020.
Citation: Kartika, K., Lakitan, B., Ria, R.P., 2021. Hydro- and osmo-priming effects on upland rice exposed to drought conditions at vegetative and reproductive stages. CMUJ. Nat. Sci. 20(3): e2021053.
INTRODUCTION
Rice cultivation at irrigated paddy fields has been well established with high productivity. Meanwhile, average productivity of upland rice is only about half of irrigated rice. Expanding paddy fields for rice production is costly, required major infrastructure development, and not always be ecologically and/or economically feasible. However, massive cultivation of upland rice by large number of smallholder farmers could collectively contribute to efforts in increasing food availability for continuously rising population in developing countries.
Natural constraint in upland rice cultivation is reliable sources of water. Water is essential for seed germination, vegetative growth stage, and especially during booting stage to few days before harvest. Massive yield loss is mostly attributed to drought occurrence during seed germination and reproductive organ development.
Drought inhibited some plant physiological and biochemical processes which, in turn, restraining plants growth and development (Larkunthod et al., 2018; Kartika et al., 2020; Lakitan et al., 2020). Drought stress caused disruption of plant metabolic processes, including nutrient absorption (da Silva et al., 2011), cell division and enlargement (Avramova et al., 2015), enzyme activity (Abedi and Pakniyat, 2010), and stomatal conductance (Kartika et al., 2020). Stomatal closure is the first reaction of plants in water deficient condition which inhibited gas exchange, including photosynthesis, transpiration, and related carbohydrates metabolism (Pirasteh- Anosheh et al., 2016). Alteration in plant physiological and biochemical under drought stress condition resulting in yield reduction. Besides affecting quantitative yield, drought also lowered grain quality in rice, including grain shape, percentage of brown rice, chalkiness, and nutrient content (Yang et al., 2019).
Seed priming has been used for enhancing germination and improving seedling quality (Ria et al., 2020). Seed priming stimulated metabolic process at pre-germinative stage but disrupted actual radicle and plumule emergence (Raj and Raj, 2019). Recently, there are many priming techniques, i.e. hydro-, osmo-, halo-, matrix-, hormone-, and nutria-priming. However, hydro- and osmo-priming are affordable, simple, and more likely to be adopted by smallholder farmers. Hydro-priming utilizes pure water, while osmo-priming uses chemical solution. Seed priming process was managed for specific time under controlled condition and was dehydrated and stored until it was ready to be planted (Pawar and Laware, 2018). Both hydro- and osmo- priming had been reported to have positive impact for improving germination (Ghassemi-Golezani et al., 2008; Lemmens et al., 2019) and seedling growth (Oliveira et al., 2019).
Although the main effect is an acceleration of germination, priming treatment has also been reported to improve vegetative growth and grain yield in rice (Mahajan et al., 2011) and ability of rice crop to tolerate drought condition (Hussain et al., 2017). However, response among rice varieties might be different to hydro- and osmo-priming treatments. Therefore, this research was conducted to evaluate effectiveness of hydro- and osmo-priming on seed germination, vegetative growth and yield of Situ Patenggang, an upland rice variety, exposed to drought stress during late vegetative, booting, and heading stages.
MATERIALS AND METHODS
Seed material and priming procedure
Upland indica rice studied was Situ Patenggang variety. Seeds with pretested germination rate higher than 90% and initial moisture contents of 10.03% were used in this study. Seeds were treated with hydro-priming or osmo-priming with different concentrations of polyethylene glycol (PEG 6000), i.e. 10%, 15%, and 20%. Seeds treated with hydro-priming were soaked in aerated distilled water. Seeds of hydro- and osmo-priming were soaked for 12 hours in complete darkness at 28 ± 2˚C. The primed seeds were rinsed with distilled water for 3-4 times; then, air dried at 28 ± 2˚C for restoring seed moisture to the pre-treatment level.
Germination and seedling growth
Primed seeds were arranged in petri dish and soaked with distilled water for 32 hours. Germination was defined by the emergence of the radicle from the seed coat. Seed emergence were counted every 4-hour intervals from 12 until 32 hours, after no further additional seed germination was detected. Time taken to 50% germination (T50) was calculated based on the formula of Coolbear et al. (1984). Mean germination time (MGT) was calculated according to Ellis and Roberts (1981). Germination index (GI) was calculated using the formula proposed by Khan and Ungar (1998). Vigor index (VI) was calculated based on percentage of germination and seedling length at 7 days after sowing (DAS) as described by Abdul-Baki and Anderson (1973).
Formula for calculating T50 was:

Germinated seeds were sown in seedling tray filled with soil as growing substrate. Seedling shoot length was measured at 7 DAS and 14 DAS by selecting 3 seedlings randomly for each treatment. Seedling data were destructively collected at 14 DAS for measuring length as well as fresh and dry weight of shoot and roots.
Pot experiment and drought stress treatment
One seedling of two weeks old were transplanted into pot filled with soil-manure mixed growing substrate (2:1 v/v). Pots were 26 cm in height, 22 cm at base diameter and 30 cm at upper diameter. Plants were supplied with enough water at early vegetative stage. Drought stresses initiated at three different stages, i.e. late vegetative stage, started at 49 days after sowing (DAS); booting stage, started at 63 DAS; or heading stage started at 91 DAS.
Plants were placed in a plastic house to avoid unexpected rainfall during drought treatment period. Drought treatment was conducted by halting water supply and prevent the plants from rain until soil moisture was dropped to less than 10%; required 14 consecutive days during late vegetative stage, and 12 consecutive days during booting and heading stages. Soil moisture was monitored daily using soil moisture meter (Lutron PMS-714) at 10 cm depth during treatments.
Measured parameters during vegetative growth and reproductive stage included leaf area (LA), shoot length (SL), root length (RL), shoot fresh weight (SFW), root fresh weight (RFW), shoot dry weight (SDW), root dry weight (RDW), root length (RL), leaf rolling score (LRS), number of tillers (NoT), time of physiological maturity (TPM), panicle length (PL), weight of 100 grains (W100), number of spikelet (NoS), and yield. LRS was observed during last two days of drought stress according to the Standard Evaluation System for Rice (IRRI, 2002). At termination of drought treatment, all plants were re- arranged to their original positions at the open field area and generously re-watered until harvest.
Experimental design and statistical analysis
This study was arranged in a randomized block design for germination and seedling growth; but, factorial randomized complete block design for pot experiment. Observed variables were measured with three replications. Each hill is assigned as replication. Collected data were subjected to ANOVA using SAS 9.0 statistical analysis software (SAS Institute Inc., Cary, North Carolina, US). Significant differences between treatments were tested using the least significant difference (LSD) test at P ≤ 0.05.
RESULTS
Effect of seed priming on germination and early vegetative growth
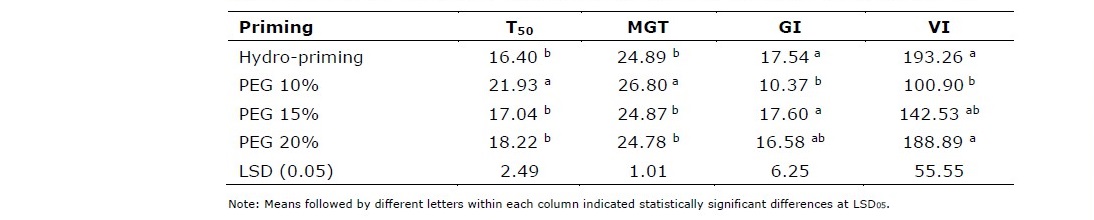
The application of hydro- and osmo-priming with application rate up to 20% did not indicate significant effect on percentage of seed emergence (Table 1). However, mean comparison of priming treatments for T50 revealed that seeds treated with 10% PEG was significantly slower to germinate than those treated with hydropriming or osmo-priming at higher PEG treatments. The application of osmo-priming decreases osmotic potential and inhibits imbibition process and metabolic activities. The application of PEG 10% took 21.93 hours to reach T50 with MGT at 26.80 hours. Furthermore, GI and VI of seeds treated with PEG 10% were 10.37 and 100.90, respectively. These values were also lower than those treated with higher PEG concentrations (Table 2). This results indicated that hydropriming was better for stimulating seed germination of Situ Patenggang variety. Otherwise, if osmo-priming used, higher concentrations (15% or 20%) of PEG was required.
Table 1. Emergence percentage of hydro and PEG-primed seeds counted every 4 hours after germination initiation.
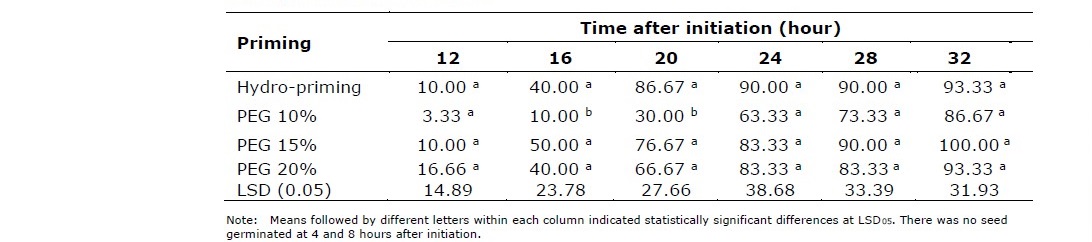
Table 2. Time taken to 50% germination (T50), means germination time (MGT), germination index (GI), and vigor index (VI) of hydro and PEG-primed seeds.
Both shoot and root growth in all priming treatments were not significantly different as indicated by their length and weight, except in leaf area and root fresh weight. Interestingly, leaf area and root fresh weight of seedlings treated with PEG 10% was the highest amongst hydro or osmo-primed seedlings at higher PEG concentrations (Table 3).
Table 3. Effects of seed priming on rice seedlings growth at 14 days after sowing.
During vegetative stage at 5 WAT and 6 WAT, PEG-priming at concentration of 15% produced significantly taller plants than hydro and other osmo-priming treatments. However, the differences in plant height were diminished at 7 WAT. This phenomenon suggested that at 7 WAT height of Situ Patenggang variety had reached its near maximum height and vegetative growth progressively shifted from increasing height to developing tillers. Rice plant has been well known for having a maximum height. However, number of tillers per hill may variably depending on genetics, environmental conditions, and cultivation practices. In this study, highest number of tillers produced by rice plants treated with hydropriming (Figure 1). However, it should be kept in mind that not all tillers were productive.
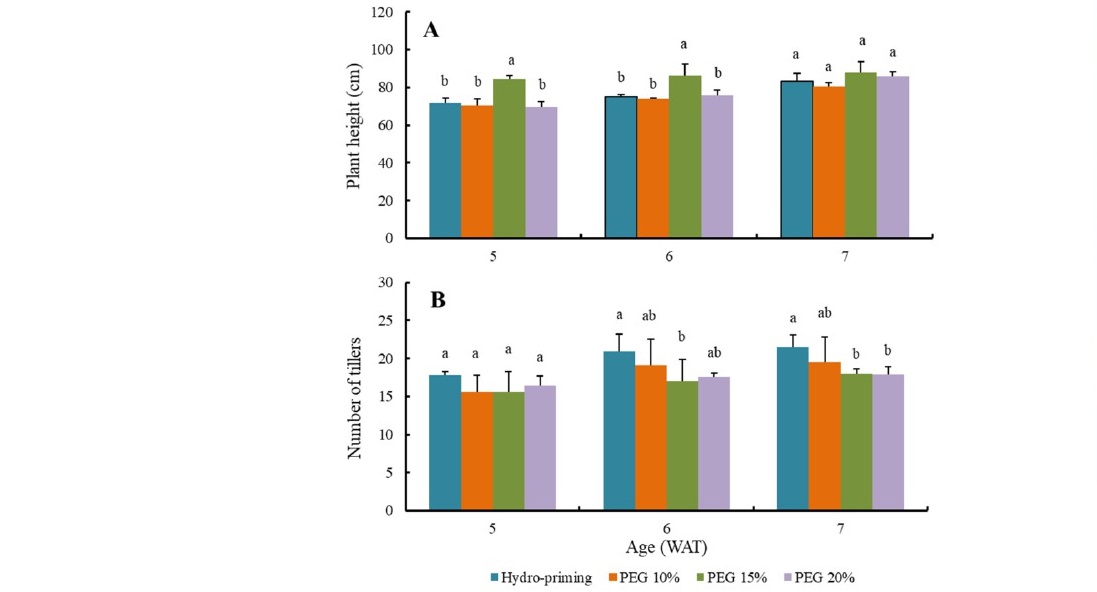
Figure 1. Effect of seed priming on plant height (A) and number of tillers (B) at 5, 6, and 7 week after transplanting.
Effect of seed priming and drought stress on growth and yield components
Drought stress was initiated at 3 different growth stages i.e. late vegetative, booting, and heading stage. Drought stress indicated by decline of water availability in the soil. The amount of water absorbed by plants was higher during generative phase including booting and heading stages. Thus, soil moisture decreased faster at generative phase. The application of different seed priming methods did not affect soil moisture at booting and heading stages. Furthermore, leaf rolling score did not affected by seed priming, but the score was higher if plant imposed to drought stress during generative phase (Figure 2).
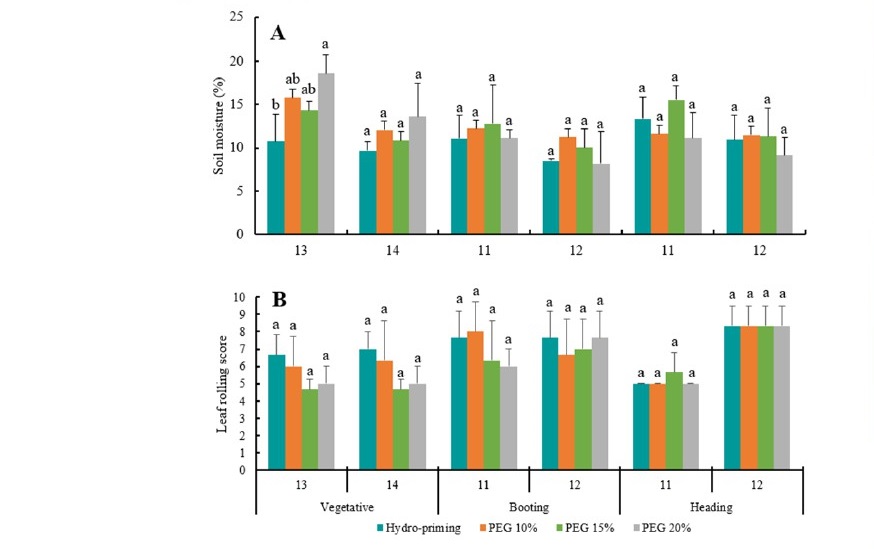
Figure 2. Moisture of growing substrate (A) and leaf rolling score (B) at the last two days of drought stress during vegetative, booting, and heading stage.
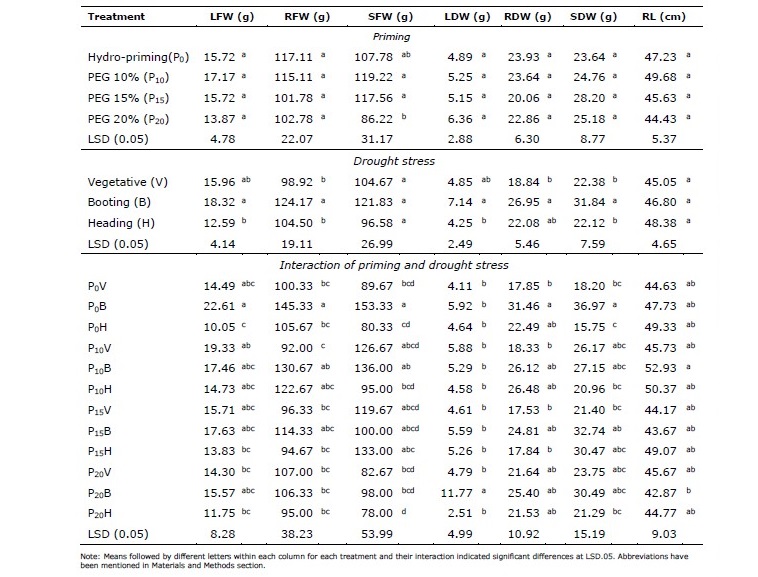
Application of seed priming did not significantly affect plant biomass except for stem fresh weight (Table 4). Plant treated with PEG 10% and PEG 15% exhibited heavier stem fresh weight of 119.22 g and 117.56 g, respectively. Plants imposed to drought stress during booting stage showed the highest weight of leaf, root, and stem. However, plant biomass significantly decreased when drought stress occurred during heading stage. Overall, the highest plant biomass obtained in plants treated with hydro- priming and experienced less stress during heading stage. Longest root was obtained by plants primed with 10% PEG priming. Plants treated with PEG10% were more able to endure drought stress during heading stage, as indicated by significant root length compared to plants treated with other priming procedures.
Table 4. Effects of seed priming, drought stress, and their interactions on the growth of vegetative organs.
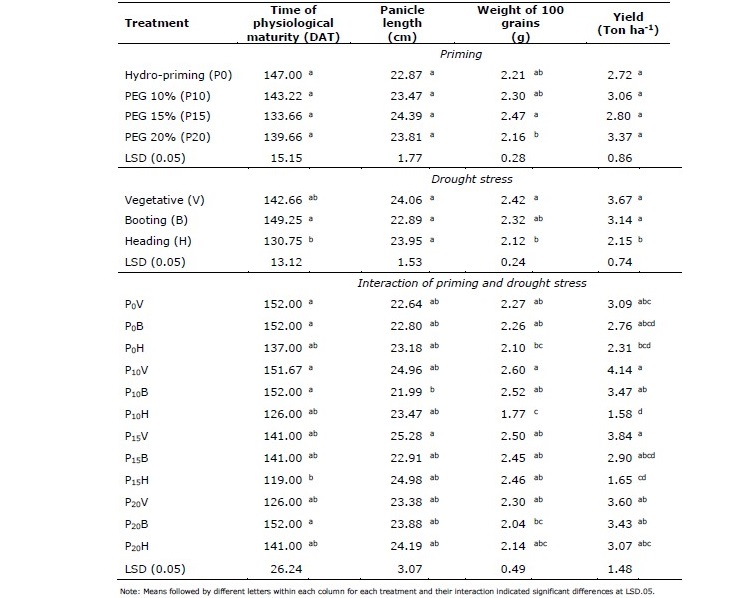
The effects of seed priming were visible in rice generative traits (Table 5). Hydro- and osmo-priming with PEG application rate up to 15% resulted heavier grains. Meanwhile, high concentration of PEG application at 20% decreased grain size as indicated by lowest weight of 100 grains. Grain weight was reduced by drought exposure. The reduction was higher if drought was imposed at generative stage. Drought stress at vegetative stage did not affect grain weight and number of filled spikelet regardless of variable priming treatments. Drought stress during late vegetative stage and booting stage decelerated maturity process and grain filling as indicated by smaller grains. Moreover, plants treated with drought stress at heading stage shortened life cycle of rice plants as indicated by earlier time of physiological maturity yet produced lower yield (Table 5).
Table 5. Effects of seed priming, different time of drought exposure, and interactions between priming and drought on yield components.
The highest number of panicles collected from three consecutive harvests (Harvest 1, 2, and 3) was found in rice plant pre-treated with hydropriming and exposed to drought at booting stage. Meanwhile, the lowest was found in plant pre-treated with PEG 15% and exposed to drought at heading stage. Conversely, percentage of filled spikelet was the lowest in the plant exhibited the highest number of panicles (Figure 3).
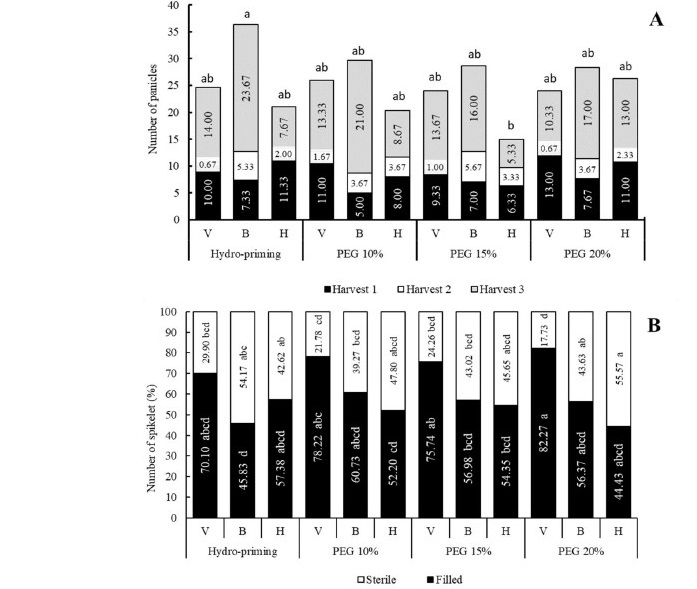
Figure 3. Interaction between seed priming and drought stress at different growth stages for number of panicles (A) and percentage of filled spikelet (B).
DISCUSSION
Seed priming on germination and seedling growth
Seed priming was implemented to improve germination rate, seedling quality which determines growth and yield of plant. The present study compared hydro-priming with 3 different concentrations of PEG used in osmo-priming treatments. Results of the study revealed that seed priming treatments caused different effects on germination, including time to 50% germination, mean germination time, germination index, and vigor index. Germination of Situ Patenggang rice variety treated with hydro-priming was comparable with osmo-priming with the application rate at 15% and 20% PEG; meanwhile, germination and vigor indexes of seeds treated with 10% PEG were significantly lower (Table 1 and 2). Similar result was previously reported in Batu Tegi, another Indonesian upland rice variety (Abdallah et al., 2016). Singh et al. (2015) recognized that effects of seed priming might be varied as affected by some factors, including specific germplasm of plant species, growing media, kind and concentration of chemical used, duration of priming treatment, and seed storage conditions after priming treatment.
Prior to germination, imbibition and activation phases occurred. Water quickly uptake during imbibition phase and causing activation of metabolic and physiological activities (Pawar and Laware, 2018). Hydro- and osmo-priming stimulates those phases but prevent radicle protrusion. Immediate germination occurs once seeds are sown. Enhanced germination is associated with several process during priming treatment, including enzyme activation, synthesis of primary metabolites i.e. proteins, nucleic acids, and ATP (Oliveira et al., 2019). The physiological changes increased starch hydrolysis and sugars availability for enhancing germination and seedling growth (Hussain et al., 2017).
Growth of seedlings treated with hydro-priming and osmo-priming at 15% and 20% PEG was not significantly different. Interestingly, seeds primed with 10% PEG exhibited better seedling growth than other priming treatments, as indicated by higher leaf area and root fresh weight (Table 3). This result was in line with report of Faijunnahar et al. (2017) that the best seedling growth was obtained when wheat seed was treated with 10% PEG compared to non-priming, hydropriming, and seedling growth gradually decreased in higher PEG concentrations. The suppression of seedling growth at high PEG concentrations has been reported as due to nutrient imbalance, mobility of toxic ions, and reduction in solubility of applied solutes (Abiri et al., 2016).
Rice seedling predominantly uses available nutritions in cotyledon during their early growth stage. At later stage, seedling growth depends on ability of roots in absorbing water and soil nutrients and harvesting light energy by newly developed leaves via photosynthesis (Artadana et al., 2019). As measured at 14 DAS, higher leaf area and longer roots of seedlings primed with 10% PEG are beneficial for supporting growth and development. Seedling growth was completely dependent on its roots for absorbing water and nutrients and leaves for capturing CO2 and light energy, since cotyledon had been fallen off the mesocotyl.
Carry-over effects of seed priming on vegetative growth
The effect of priming treatment on rice vegetative growth was mainly exhibited on development of tillers. Number of tillers is important since it determines number of panicles produced and potential yield in rice crop (Kartika et al., 2018a). Higher number of tillers was produced in hydro-priming treatment compared to osmo-priming with 10% to 20% PEG concentrations (Figure 1). Tillers initiation seemed to be triggered by drier growing substrate, since moisture dropped to a much lower level (around 10%) in the substrate grown with hydro-primed rice plant; meanwhile, all recorded moisture levels were consistently higher in substrate grown with osmo-primed plants, varied from 10% to 20% PEG concentrations (Figure 2). Alam et al. (2015) mentioned some genes that could contribute to increase of photosynthesis and tiller numbers in rice plants exposed to salinity and drought conditions. Further research is needed to prove whether Situ Patenggang variety also contains the drought tolerant genes.
All rice plants exposed to 14 days drought treatments reached maximum leaf rolling index regardless of seed priming treatment selected if drought was exposed at heading stage (Figure 2). Leaf rolling index has been used for visual indicator for drought stress in rice plant (Kartika et al., 2018b). As for plant biomass, higher stem fresh weight identified in 10% and 15% of PEG priming application and less reduction in biomass of vegetative organs was observed if drought was exposed during booting stage (Table 4). Drought during booting stage severely suppressed newly developed water- sensitive inflorescence. This might cause relocation of assimilates produced by photosynthesis to further support growth of vegetative organs. Kobata et al. (1994) reported that drought during booting stage did not reduce assimilate production but did increase sterility in spikelet. These findings support the possibility of assimilate relocation to vegetative organs since the damaged reproductive organ lose its role as a dominant sink.
Drought effects on yield
Larger grain size was obtained in rice plant grown using seeds pre-treated with 15% PEG; however, the plant did not produce higher yield (Table 5). This indicated that the plant produced lower total grains per hill than plants treated with other priming treatments. In this case, low total grains were consistently associated with lower number of tillers developed during vegetative stage, as shown at Figure 1. Despite showing positive effects on germination and early stage of seedling growth, the effects of osmo-priming did not go beyond four weeks after seedlings were transplanted. Instead, drought exposure during late vegetative stage restrained number of tillers produced per hill (Ria et al., 2020). Kang et al. (2017) had identified specific gene related to tiller development in drought tolerant rice. During recovery after drought treatment was terminated, assimilates were mainly allocated for supporting development of reproductive organs within the limited number of tillers, resulting larger grain size.
Among seed-primed plants exposed to drought, the highest yield was attained in plants pre-treated with 10% PEG and exposed to drought only during vegetative stage. In this study, panicle length, grain size, and yield in rice plants exposed to drought during vegetative stage were less severely affected compared to those exposed to drought during booting or heading stage. Similar results were reported by Zhang et al. (2018) who found that drought had larger detrimental influences during the reproductive phase, e.g., blooming stage, filling stage, and maturity.
Drought exposure during heading stage enhanced maturity but produced less yield (Figure 5). Drought stress reduced endogenous auxins production but increased abscisic acid and ethylene, which in turn, fastened flowering and grain maturity (Abobatta, 2019). Shortened time to physiological maturity potentially reduced grain yield due to short available time for accumulation of photosynthates (Shavrukov et al., 2017).
Plant biomass and yield components were more significantly affected by drought stress condition. As rice plants shifted from vegetative to generative stage, the plants had reached maximum vegetative organ growth. Situ Patenggang rice variety was able to maintain their yield if drought stress occurred during vegetative stage. Similar results reported in another upland rice varieties i.e Inpago 4, Inpago 10, and Rindang 1 (Ria et al., 2020). Reduction of grain weight and number of filled spikelet occurred when plant imposed to drought stress during generative stage. Farooq et al. (2009) reported that yield reduction under drought stress was due to reduction of photosynthetic rate. Akram et al. (2013) estimated that drought stress reduced photosynthetic rate by 30.69% at panicle initiation, 28.00% at anthesis, and 12.26% at grain filling. In addition, reproductive abortion also happened as the capacity of sinks to utilize assimilates declined.
CONCLUSION
Seed priming exhibited positive effects on germination and early growth of rice seedlings. The effects of seed priming gradually diminished soon after the plants were exposed to drought at late vegetative stage. Development of reproductive organs were predominantly regulated by drought during booting and heading stages. Drought during late vegetative stage suppressed tillers development but the plants were able to recover and produced comparable yield with their significantly higher percentage of filled spikelet and larger grain. Rice plants exposed to drought at booting stage were inherited with higher number of tillers per hill since the plants had produced optimum number of tillers before drought exposure; however, they produced higher percentage of sterile spikelet. Physiological maturity in rice plants was enhanced by drought exposure during heading stage but this survival mechanism was at cost of yield reduction.
ACKNOWLEDGEMENTS
Authors of this article would like to thank the editor of this journal and the anonymous reviewers for their comments and suggestions.
REFERENCES
Abdallah, E.H., Musa, Y., Mustafa, M., Sjahril, R., and Riadi, M. 2016. Comparison between hydro and osmo-priming to determine period needed for priming indicator and its effect on germination percentage of aerobic rice cultivars (Oryza Sativa L.). Agrivita. 38: 222–230.
Abdul‐Baki, A.A., and Anderson, J.D. 1973. Vigor Determination in Soybean Seed by Multiple Criteria 1. Crop Science. 3: 630–637.
Abedi, T., and Pakniyat, H. 2010. Antioxidant enzyme changes in response to drought stress in ten cultivars of oilseed rape (Brassica napus L.). Czech Journal of Genetics and Plant Breeding. 46: 27–34.
Abiri, R., Shaharuddin, N.A., Maziah, M., Yusof, Z.N.B., Atabaki, N., Sahebi, and M., Azizi, P., 2016. Quantitative assessment of Indica rice germination to hydropriming, hormonal priming and polyethylene glycol priming. Chilean Journal of Agricultural Research. 76: 392–400.
Abobatta, W.F. 2019. Drought adaptive mechanisms of plants – a review. Advances in Agriculture and Environmental Science: Open Access (AAEOA). 2: 42–45.
Akram, H.M., Ali, A., Sattar, A., Rehman, H.S.U., and Bibi, A. 2013. Impact of water deficit stress on various physiological and agronomic traits of three Basmati rice (Oryza sativa L.) cultivars. Journal of Animal and Plant Sciences. 23: 1415–1423.
Alam, M.M., Tanaka, T., Nakamura, H., Ichikawa, H., Kobayashi, K., Yaeno, T., Yamaoka, N., Shimomoto, K., Takayama, K., Nishina, H. and Nishiguchi, M., 2015. Overexpression of a rice heme activator protein gene (Os HAP 2E) confers resistance to pathogens, salinity and drought, and increases photosynthesis and tiller number. Plant Biotechnology Journal, 13: 85-96.
Artadana, I.B.M., Dewi, I.T., and Sukweenadhi, J. 2019. The performance of three local rice (Oryza sativa L.) cultivar from East Kalimantan-Indonesia under drought stress at early seedling stage, in: IOP Conference Series: Earth and Environmental Science. 1–8.
Avramova, V., Abdelgawad, H., Zhang, Z., Fotschki, B., Casadevall, R., Vergauwen, L., Knapen, D., Taleisnik, E., Guisez, Y., Asard, H., and Beemster, G.T.S. 2015. Drought induces distinct growth response, protection, and recovery mechanisms in the maize leaf growth zone. Plant Physiology. 169: 1382–1396.
Coolbear, P., Francis, A., and Grierson, D. 1984. The effect of low temperature pre- sowing treatment on the germination performance and membrane integrity of artificially aged tomato seeds. Journal of Experimental Botany. 35: 1609–1617.
da Silva, E., Nogueira, R., da Silva, M., and de Albuquerque, M. 2011. Drought stress and plant nutrition. Plant Stress 5: 32–41.
Ellis, R.H., and Roberts, E.H. 1981. The quantification of ageing and survival in orthodox seeds. Seed Science and Technology. 9: 373–409.
Faijunnahar, M., Baque, A., Habib, M.A., and Hossain, H.M.M.T. 2017. Polyethylene glycol (PEG) induced changes in germination, seedling growth and water relation behavior of wheat (Triticum aestivum L.) genotypes. Universal Journal of Plant Science. 5: 49–57.
Farooq, M., Wahid, A., Kobayashi, N., Fujita, D., and Basra, S.M.A. 2009. Plant drought stress: Effects, mechanisms and management. Sustainable Agriculture. 29: 185–212.
Ghassemi-Golezani, K., Aliloo, A.A., Valizadeh, M., and Moghaddam, M. 2008. Effects of different priming techniques on seed invigoration and seedling establishment of lentil (Lens culinaris Medik). Journal of Food, Agriculture and Environment. 36: 29–33.
Hussain, M., Farooq, M., and Lee, D.J. 2017. Evaluating the role of seed priming in improving drought tolerance of pigmented and non-pigmented rice. Journal of Agronomy and Crop Science. 203: 269–276.
IRRI. 2002. Standard evaluation system for rice. International Rice Research Institute, Los Banos, the Philippine.
Kang, J., Li, J., Gao, S., Tian, C., and Zha, X. 2017. Overexpression of the leucine‐rich receptor‐like kinase gene LRK 2 increases drought tolerance and tiller number in rice. Plant biotechnology journal. 15: 1175-1185.
Kartika, K., Lakitan, B., Sanjaya, N., Wijaya, A., Kadir, S., Kurnianingsih, A., Widuri, L.I., Siaga, A., and Meihana, M. 2018a, Internal versus edge row comparison in jajar legowo 4:1 rice planting pattern at different frequency of fertilizer applications. AGRIVITA Journal of Agricultural Science, 40: 222–232.
Kartika, K., Lakitan, B., Wijaya, A., Kadir, S., Widuri, L.I., Erna Siaga, E., and Meihana, 2018b, Effects of particle size and application rate of rice husk biochar on chemical properties of tropical wetland soil, rice growth, and yield. Australian Journal of Crop Science. 12: 817-826.
Kartika, K., Sakagami, J.I., Lakitan, B., Yabuta, S., Wijaya, A., Kadir, S., Widuri, L.I., Siaga, E., and Nakao, Y. 2020. Morpho-Physiological Response of Oryza glaberrima to Gradual Soil Drying. Rice Science. 27: 67–74.
Khan, M.A., and Ungar, I.A. 1998. Germination of the salt tolerant shrub Suaeda fruticosa from Pakistan: Salinity and temperature responses. Seed Science and Technology. 26: 657–667.
Kobata, T., Tanaka, S., Utumi, M., Hara, S. and Imaki, T., 1994. Sterility in rice (Oryza sativa L.) subject to drought during the booting stage occurs not because of lack of assimilate or of water deficit in the shoot but because of dehydration of the root zone. Japanese journal of crop science, 63: 510-517.
Lakitan, B., Jaya, K.K., Ria, R.P., and Morianto, B. 2020. The Effects of Different NPK Fertilization Rates and Water Regimes on Ratooned Black Glutinous Rice. CMU Journal of Natural Science, 19: 350-365.
Larkunthod, P., Nounjan, N., Siangliw, J.L., Toojinda, T., Sanitchon, J., Jongdee, B., and Theerakulpisut, P. 2018. Physiological responses under drought stress of improved drought- Tolerant rice lines and their parents. Notulae Botanicae Horti Agrobotanici Cluj-Napoca. 46: 679–687.
Lemmens, E., Deleu, L.J., De Brier, N., De Man, W.L., De Proft, M., Prinsen, E., and Delcour, J.A., 2019. The impact of Hydro-priming and Osmo-priming on seedling characteristics, plant hormone concentrations, activity of selected hydrolytic enzymes, and cell wall and phytate hydrolysis in sprouted wheat (Triticum aestivum L.). ACS Omega 4: 22089–22100.
Mahajan, G., Sarlach, R.S., Japinder, S., and Gill, M.S. 2011. Seed priming effects on germination, growth and yield of dry direct-seeded rice. Journal of Crop Improvement. 25: 409–417.
Oliveira, C.E.D.S., Steiner, F., Zuffo, A.M., Zoz, T., Alves, C.Z., and De Aguiar, V.C.B. 2019. Seed priming improves the germination and growth rate of melon seedlings under saline stress. Ciencia Rural. 49: 1–11.
Pawar, V.A., and Laware, S.L. 2018. Seed Priming A Critical Review. International Journal of Scientific Research in Biological Sciences. 5: 94–101.
Pirasteh-Anosheh, H., Saed-Moucheshi, A., Pakniyat, H., and Pessarakli, M. 2016. Stomatal responses to drought stress, in: Water Stress and Crop Plants: A Sustainable Approach. (pp. 24–40). John Wiley & Sons, Ltd., Hoboken.
Raj, A.B., and Raj, S.K. 2019. Seed priming: An approach towards agricultural sustainability. Journal of Applied and Natural Science. 11: 227–234.
Ria, R.P., Lakitan, B., Sulaiman, F., Kartika, K. and Suwignyo, RA. 2020. Cross- ecosystem utilizing primed seeds of upland rice varieties for enriching crop diversity at riparian wetland during dry season. Biodiversitas. 21: 3008–3017.
Shavrukov, Y., Kurishbayev, A., Jatayev, S., Shvidchenko, V., Zotova, L., Koekemoer, F., De Groot, S., Soole, K., and Langridge, P. 2017. Early flowering as a drought escape mechanism in plants: How can it aid wheat production? Frontiers in Plant Science. 8: 1950.
Singh, H., Jassal, R.K., Kang, J.S., Sandhu, S.S., Kang, H., and Grewal, K. 2015. Seed priming techniques in field crops -A review. Agricultural Reviews. 36: 251–264.
Yang, X., Wang, B., Chen, L., Li, P., and Cao, C. 2019. The different influences of drought stress at the flowering stage on rice physiological traits, grain yield, and quality. Scientific Reports. 9: 3742.
Zhang, J., Zhang, S., Cheng, M., Jiang, H., Zhang, X., Peng, C., Lu.,X., Zhang, M., and Jin, J. 2018. Effect of drought on agronomic traits of rice and wheat: a meta- analysis. International Journal of Environmental Research and Public Health 15: 839.
OPEN access freely available online
Chiang Mai University Journal of Natural Sciences [ISSN 16851994]
Chiang Mai University, Thailand https://cmuj.cmu.ac.th
Kartika Kartika1,2, Benyamin Lakitan1,3,*, Rofiqoh Purnama Ria1
1 College of Agriculture, Universitas Sriwijaya, Inderalaya 30662, Indonesia
2 Research Center for Biology, Indonesian Institute of Science (LIPI), Cibinong 16911, Indonesia
3 Research Center for Sub-optimal Lands (PUR-PLSO), Universitas Sriwijaya, Palembang 30139, Indonesia
Corresponding author: Benyamin Lakitan, E-mail: blakitan60@unsri.ac.id
Total Article Views
Editor: Wasu Pathom-aree,
Chiang Mai University, Thailand
Article history:
Received: August 14, 2020;
Revised: December 6, 2020;
Accepted: January 15, 2021;
Published online: March 15, 2021